SDG 14: Life Below Water

SDG 14: Life below water. Conserve and sustainably use the oceans, seas and marine resources for sustainable development
Introduction
Our planet is blue. Seas and oceans cover more than 70% of the Earth’s surface. Although humans are terrestrial organisms – we live on the continents, like on large islands – our existence is closely tied to the water on our planet. Oceans provide us with water, food, energy, and a range of natural resources. They are used as trade and communication routes. Apart from these, they serve as climate and weather regulators by acting in the global water cycle. They provide environments for the countless biodiversity of living creatures – animals, plants, fungi, and microorganisms – ranging from sunny beaches to dark and cold deeps.
Researchers face challenges while exploring life in water, partly due to the water’s physical characteristics – e.g., its density, the pressure that increases with depth, along with light absorption and scattering (which vary by wavelength) that complicate direct observations. Fortunately, growing scientific and technical abilities enable us to investigate life in oceans better than ever in our history, allowing us to learn about processes hidden in the waters that influence our existence on our planet. Also, to estimate how the impact of human activity changes the water environment, and what consequences will come back to us. Healthy oceans, and, in general, a healthy hydrosphere, are crucial for our survival on Earth. By studying the “Life Below Water” chapter, you will become an environmentally responsible citizen of our planet.
1. Brief (hi)story of Earth and water
The Earth, the third planet in the Solar System, is known as a “Blue Planet1”. Although its surface contains gases of the atmosphere, rocks of the lithosphere (both continental and oceanic crusts), and water of the hydrosphere, the latter one in the form of oceans, cover more than 70% of our planet (e.g., NOAA 2025a). The oceans also contain the vast majority of the total planet’s liquid water (about 1,338,000,000 km3 – thus more than 95% of the total 1,386,000,000 km3) and provide the primary source of water for evaporation, condensation in clouds, and future precipitation in rain, playing important role in so-called water cycle (e.g., Duxbury, Duxbury & Sverdrup 2002; Meran, Siehlow & von Hirschhausen 2021; Shiklomanov 1990). Nevertheless, the mass of ocean water is only a minute fraction of the total mass of our planet (about 0.02 %), and it covers only a skinny surface layer of the uppermost part of the planet’s lithosphere, the crust. The average depth of all oceans is only 3,682 m. The deepest site, Challenger Deep, discovered within the Mariana Trench (Pacific), reaches so much, and still only about 10,925 meters below the ocean’s surface (Genda & Ikoma 2008; NOAA 2024a; Stewart & Jamieson 2019). This means there is an average of just 3.7 km of water and, in maximum, almost 11 km of water on the surface of the sphere with a radius of nearly 6,400 km (Mamajek et al. 2015).
Why is there water on Earth?
The question of the origin of water on Earth is not easy to resolve. Earth’s age is estimated to be 4.55 Ga (e.g., Allègre, Manhès & Göpel 1995; NASA 2025). Evidence from ancient zircon isotopes indicates the presence of liquid water interacting with the crust by 4.3 Ga (Mojzsis, Harrison & Pidgeon 2001; Pinti 2005), which later created conditions favorable for the evolution of life (evidenced already by fossils of microbial structures age 3.7 Ga; Dodd et al. 2017; Nutman et al. 2016). Its presence can be explained by an internal (indigenous origin) model, resulting from the planet’s formation processes that trapped the water in mantle minerals (if Earth’s orbit allows hydrous phases to be stable). But because of its location in the early Solar System, the early Earth was believed to reach too high temperatures to support the water in the hydrous phase. Therefore, the exogenous explanation, which includes comets and asteroids (chondrite meteorites), which are now believed to be the main source of water, is a viable way to supply water to the Earth after it had formed (Drake 2005; Marty 2012; Meech & Raymond 2019; Piani et al. 2020; Sarafian et al. 2017). A current consensus interpretation assumes a combination of sources rather than a single source from those listed above (with the majority delivered by meteorites; Wu et al. 2018). Water could then be released through e.g., volcanic activity, the outgassing (e.g. Drake 2005; Grott et al. 2011) to become a part of the initial atmosphere, and when the temperatures on Earth’s surface dropped, the water vapor condensed and formed the first liquid oceans (Abe 1993; Abe & Matsui 1985; Mojzsis, Harrison & Pidgeon 2001; Pinti 2005).
The proper position of Earth’s orbit in the so-called habitable zone around our Sun (the appropriate distance) ensures the presence of liquid water on the surface (Kasting, Whitmire & Reynolds 1993). Together with the proper position, the Earth’s rotation and atmosphere composition (especially the heat-trapping gases like water vapor, carbon dioxide, or methane responsible for a so-called greenhouse effect2) ensure the temperatures on the surface are warm enough to sustain the oceans in their liquid state (Duxbury, Duxbury & Sverdrup 2002; Smith 2008). Although the volume of water on Earth remains constant, the thermal energy of the Sun, combined with the Earth’s gravity, causes the continuous movement of water particles, thereby ensuring the global water cycle.
How is water cycling on Earth?
The water on Earth is in continuous motion. It circulates naturally mainly due to the energy of solar radiation, which leads to evaporation from the surface of the oceans and other water sources. Liquid water changes into vapor, forms clouds, and then it may recondense back into liquid (and gravity drives its downward flow). It may happen in the same place or elsewhere, due to the wind transportation in the atmosphere (e.g. Perlman & Evans 2019; Fig. 1). According to Oki and Kanae (2006), nearly 90% of the water that evaporates from the oceans, precipitates back into them, and only a small fraction is transported in the atmosphere over terrestrial areas. The water from the rain lacks sea salt, so it is freshwater. It soaks the ground, infiltrates into the groundwater, fills lakes, wetlands, and human-made reservoirs (e.g., dam pools, agricultural irrigations). Where temperatures are low, it forms permafrost, snow and ice, or glaciers that eventually melt. The water surplus runs off on the terrain surface, and in the streams and rivers, it comes back to the seas and oceans. During all these processes, it evaporates back into the atmosphere too (Oki & Kanae 2006; USGS 2025). Part of the water is temporarily trapped in bodies of living organisms (which can live in any of the above-listed environments). The process is called the water cycle or hydrological cycle and plays an important role in climate regulation on our planet (e.g. the water vapor is a greenhouse gas in the atmosphere and may act in a positive feedback loop: the higher the temperature is, the more evaporation occurs, so more vapor is in the atmosphere and temperature rises more…; e.g. Trenberth 2011). Oki and Kanae (2006) expect the climate change to accelerate the global hydrological cycle with the growth of precipitation worldwide.
2. Water as an environment for life
What is water, in the context of life?
Humans are terrestrial and air-breathing organisms; this fact poses a challenge to direct observations. The anthropogenic attempt to observe life below water is biased by the biology of an observer. This implies that our understanding of what happens below the water surface still is quite unsatisfying, and such research requires tools that allow deep-sea investigations – which were developed relatively recently in our history (e.g. Barber and Hilting 2000). Despite this, we use water’s properties to define temperature in everyday life: in the Celsius scale, historically, 0 °C corresponds to the melting point of ice (at standard atmospheric pressure).
First, water is essential for the survival of all known living beings, serving as both the background matrix and an actor in cell and organism physiology, biology, and ecology (Chaplin 2006). Many of these creatures live in water, as in their environment. Oceans, seas, and freshwater environments provide diverse habitats that can be summarized into three general types: shallow bottoms with good daily light access (littoral or shallow shelf area), vast open waters (pelagic), and the deep, dark, and cold bottoms of reservoirs (benthic; to be explained later in this chapter). Organisms inhabiting these environments can be grouped into three main categories: plankton – those drifting with masses of water (like phytoplankton, krill, or some larvae of sessile species); nekton – actively swimming organisms found both in shallow and pelagic waters (including fish, mammals and mollusks); and benthic – diverse organisms associated to seafloor environments (either sessile or bottom-dwelling, ranging from cnidarians like corals, to echinoderms and mollusks) – they are present in all variables and depths of bottoms in the ocean (e.g. Cox, Moore & Ladle 2016; Duxbury, Duxbury & Sverdrup 2002).
Life evolved in water. The majority known evolutionary lineages of living organisms today, including prokaryotes (a broad group of microbes: bacteria and archaea) and eukaryotes: protists, plants, fungi, and phyla of animals, have their representatives living in water (e.g., see tree of life in Brusca, Moore & Shuster 2016; Paps et al. 2023). Nevertheless, even if majority of known phyla and classes contain marine creatures, when considering the macroscopic organisms only, just about 15%3 of known, named species live in marine environments, with some groups like echinoderms, cephalopods, or corals that remained exclusively in seawaters (see summary of distribution of biodiversity among the oceans, lands, and freshwater habitats in Grosberg, Vermeij & Wainwright 2012). The diversity of species decreases with depth of the water and with latitude (with distance from the tropics; Costello & Chaudhary 2017; Cox, Moore & Ladle 2016). The oceans (theoretically) provide an unlimited and large area of water. The relative homogeneity of the significantly large part of the oceans – the open waters of the pelagic zone – may explain the scarce species diversity (as habitat complexity promotes biodiversity; Leite Jardim et al. 2025). At the same time, the shallow seafloor may offer a three-dimensional landscape complexity that improves the diversity of life, with coral reefs serving as hot spots4 of marine biodiversity, hosting about one-fourth to one-third of total marine diversity (and members of 32 of 34 worldwide recognized phyla) on an area of about 1% of seafloor (NOAA 2024e; Sobha, Vibija & Fahima 2023).
Second, water on Earth exists in three states: solid (ice), liquid, and vapor. In standard atmospheric pressure conditions (1013.25 hPa), ice melts at 0 °C 5 (so it undergoes a change of state), and a liquid pure water boils at 100 °C, producing a vapor state. Each of these phase transitions requires energy (heat) to be supplied – the opposite direction is possible after energy is lost. Under Earth’s conditions, there are very few environments where water reaches 100 °C. Nevertheless, liquid water evaporates into vapor in temperatures below 100 °C – in this case, a sample of cooler water must receive more energy than a similar sample of hotter water (evaporation is a different process than boiling; it happens on the water surface when water molecules receive enough energy to be released to the air, and in the majority do not come back; see (Duxbury, Duxbury & Sverdrup 2002; USGS 2019). However, this character is fundamental for evaporation over oceans in the water cycle, or any of drying processes – from the water puddles and ponds, to the wet surfaces of organisms like moisture or our sweat from the skin, or wet feathers of some diving birds like cormorants (e.g. Baker 2019; Kadlec 2006; Rijke 1968; Srinivasan et al. 2014). Evaporated water loses heat when condensing and becomes rain. Temperatures of evaporation and freezing are, respectively, slightly higher and lower when the water is marine (salted). To warm up a sample of water, the energy must first break the intermolecular connections (bonds) between the water molecules – as a result, most of the energy is involved in this action to allow molecules in the sample to move, and the energy is released back while cooling, slowing down the molecules. This molecular mechanism underlies the feature of water called high heat capacity – meaning that when water absorbs or loses large quantities of heat, its temperature changes only slightly (it warms up slowly and cools down slowly). As a result, the temperatures of water reservoirs are much more constant than terrestrial environments in the same climatic zone. This leads to the third part – the molecular aspects of the water particle.
Third, the properties of water molecules determine the crucial physical parameters of liquid water, which are essential for life. The hydrogen-oxygen-hydrogen (H2O) bond angle is 105 °, which makes the molecule polar, which underlines the water molecule’s tendency to form dynamic clusters in the liquid state (e.g. Stillinger 1980). The sticking of molecules (by hydrogen bonding) leads to significant water features and physical anomalies. These are: large heat capacity, which, e.g., stabilizes the Earth’s climate (because water warms up slowly in summer and cools down slowly in winter), and the occurrence of maximum density at about 4 oC6 , which means that the ice is floating over the water (not freezing from bottom up, allowing the organisms to survive in deep liquid waters during winters). The same molecular property underlies the high surface tension of water, which allows some organisms to walk or live on its surface (Kontogeorgis et al. 2022). Viscosity – another result of clustering of molecules – and related features are important for life in water because it affects the active and passive movement of organisms (an actively swimming organism has a streamlined shape, while a passively moving organism may have a large surface area to remain suspended in the water column), the feeding of filter-feeding animals, as well as the spatial distribution of concentrations of various substances and temperature gradients in the environment (Duxbury, Duxbury & Sverdrup 2002; Lampert & Sommer 1996). Therefore, land (air) and water environments differ in fundamental physical properties: heat capacity, viscosity, medium density, and transparency, as well as, e.g., gases concentrations, and these have essential importance for the evolution of life and adaptations of living organisms (Grosberg, Vermeij & Wainwright 2012).
What are gradients in the water environment?
The oceans of the world are interconnected, and the water within each ocean basin slowly rotates. Hence, organisms living in the oceans or their offspring may disperse relatively easily with the movement of those waters. This explains the water-currents dependent broad distribution, in suitable habitats, of both shallow-sea species (such as oysters, mussels, or corals7) and deep-water creatures, including active and weak swimmers and sessile organisms (Babcock, Wills & Simpson 1994; Corrochano-Fraile et al. 2024; Hata et al. 2017; Lörz et al. 2023; McManus & Woodson 2012; Pineda, Hare & Sponaugle 2007; Puckett et al. 2018; Rodríguez-Flores et al. 2023; Vogt-Vincent, Mitarai & Johnson 2023). This includes extreme examples of hydrothermal vent squat lobsters that are believed to disperse as larvae with the ocean water and were found in both Pacific and Indian Ocean environments (hydrothermal, chemosynthetic-based deep-water environments are known from the high level of endemism8, reaching up to about 80% of species from this ecosystem; Hwang et al. 2022). Although there are no strict geographic boundaries within the open oceans, the depth of this environment enables the formation of gradients in physical parameters, creating different habitats. The physical properties of water, described above, underlie this vertical zonation. The most crucial for life are light, temperature, oxygen, density, biogenic nutrients, and salinity.
Light, photosynthesis, and life below the water surface
The light comes from the Sun in a predictable 24-hour cycle of Earth’s rotation. The part of light that was not reflected from the water surface is absorbed and scattered as it passes through the water column. This is why, when we look downward, we see dark waters, and when we look upward, we see brighter colors. It also explains why many aquatic animals have a dark dorsal side (to hide in the darkness below when observed from above) and a pale ventral side (to match the brightness when viewed from below; Donohue, Hemmi & Kelley 2020). The physics of light in water affects the potential for photosynthesis, since it is possible only in a visible range of the light spectrum (so-called photosynthetic available radiation [PAR]; Barber & Tran 2013; Edmondson 1956; Liu & van Iersel 2021; Marra et al. 2014). Different wavelengths of components of the visible spectrum of light penetrate to different depths in water (the red is absorbed first, then orange, and yellow, with green and blue that are available in deeper parts of the water column) – and these depths depend on water clarity9 (including the salinity or the amount of suspended biotic and abiotic particles; Pope & Fry 1997; Röttgers, McKee & Utschig 2014). The uppermost layer of waterbodies where light penetration is enough for photosynthetic organisms – both single-celled organisms called phytoplankton and benthic aquatic plants (and algae) – is called the euphotic zone10 (or productive zone). This is the zone where photosynthesis occurs, thus the primary production in the ecosystem occurs, a base for all the levels of the oceanic trophic net (the zooplankton, the fish, and, to some degree, humans too). How deep is an euphotic zone? Depending on water transparency, this is up to a few tens of meters and may vary during the year (Lampert & Sommer 1996; Lee et al. 2007; Marra et al. 2014).
The euphotic zone is a crucial environment for the development of coral reefs. These are complex, three-dimensional, and diverse environments, biologically formed in warm, shallow ocean waters. The corals – colonial sessile cnidarians from the Scleractinia order – are small jellyfish-like carnivorous organisms in the form of polyps, like tiny sea anemones or freshwater Hydra. They form a colony in which each polyp secretes a biomineral (calcium carbonate), forming a rigid skeleton that is shared with hundreds or thousands of its neighboring relatives (polyps) (Cox, Moore & Ladle 2016). This biomineral is the hard, stony base of a coral reef – that is why we call corals ecosystem engineers. Corals thus ensure a diverse structure of the environment where other organisms can live, they provide habitats and shelters, influence the availability of resources, and protect the coastline (e.g., against high-energy waves). It is estimated that coral reefs ensure an environment for about ¼ of all marine fish species, together with diverse sponges, cnidarians (including non-colonial corals), mollusks, echinoderms, crustaceans, and many more; lots of them are endemic (Burke et al. 2012; Lutzenkirchen, Duce & Bellwood 2024; Maragos, Crosby & McManus 1996). Finally, coral reefs are an important environment for humans as well. They are serving as a food source and a source of income – according to Burke et al. (2012), more than 275 million people globally live within about 10 km from the coast and may highly rely on reefs e.g., harvesting fish for food and trade, and reefs play a role as a tourism attractor. Finally, coral organisms, which produce natural venomous substances, are used to create new medicines (Bruckner 2002; Rafiudeen et al. 2024).
The reef-forming corals are called hermatypic corals. Although most inhabit semi-tropical and tropical marine environments where the temperature does not drop below 20 °C (they prefer water temperatures ranging from 23-29 °C), they can tolerate short episodes of higher temperatures; however, in general, exceeding 34 oC is dangerous for their survival (Kleypas, McManus & Menez 1999; Lough, Anderson & Hughes 2018; NOAA 2024c). They provide an extraordinary example of symbiotic interaction with a type of photosynthetic dinoflagellate algae called zooxanthellae, which live in their cells (Arabeyyat et al. 2024). In this cooperation, coral provides a safe place and nitrogenous waste products that benefit algae metabolism. In return, algae provide the energy (nourishment) derived from photosynthesis – thus, the distribution of reefs can be restricted by access to light (among other parameters). Although corals can gather their food by hunting their prey, the algae’s contribution to their survival is significant (e.g., they may exist in environments poor in nutrients, as an algae partner ensures energy; Cox, Moore & Ladle 2016; Frankowiak et al. 2016). This interaction is important enough that it may lead to the death of the coral partner if the algae die. The phenomenon called “coral bleaching” – loss of symbiotic zooxanthellae under stress (usually thermal stress, with high light) – leads to exposure of the white skeleton of the colony, because, without symbionts, the coral tissues are transparent. The bleached coral is weakened and may more easily develop disease and/or die (Brandt & McManus 2009). This was observed more commonly in the late years of the 20th and beginning of 21st century and is explained as one of the results of climate change (precisely, the increase in ocean temperature; GBRF n.d.; Lough, Anderson & Hughes 2018; Selig, Casey & Bruno 2010; Virgen-Urcelay & Donner 2023). Fortunately, if the stressor disappears quickly, the process can potentially be reversed, and symbiosis with zooxanthellae algae reappears. However, the coral remains vulnerable (e.g., the growth/reproduction rate is lower) and prone to disease (Lough, Anderson & Hughes 2018; Meesters & Bak 1993). If the stressor lasts too long, the coral dies. According to Carmignani et al. (2025), between 2009 and 2018, about 13.5% of the world’s scleractinian corals vanished. Scleractinian corals are of great interest today because they play an essential ecological role, including serving as ecological engineers, and because of the uncertain future of coral reefs due to the growing anthropogenic impact and global climate crisis (Brandl et al. 2019; Selig, Casey & Bruno 2010; Stolarski et al. 2011). Boström-Einarsson et al. (2020) emphasize the need for the restoration actions (including the role of spawning and water currents as a source of young corals for recovery). At the same time, Hughes et al. (2017) underline that the development of sustainable management of these important ecosystems – to maintain reefs and their biodiversity in stable equilibrium – requires more widely based solutions than the current emphasis on conservation, habitat protection, and restoration (e.g., to include reducing poverty actions, and encouraging shifts in human social norms-behaviors).
Coral reefs are typically considered to be distributed in shallow, warm seas. A new horizon opened with the recent advances in deep water research (technical) possibilities. The deep waters worldwide – on continental shelves, slopes, or seamounts – reveal a surprisingly diverse coral ecosystem in these dark and cold waters, also in higher latitudes. The deep-water corals (most commonly found 1-6 km below the surface) do not have a symbiotic partner (thus they do not need light), but form a long living, blooming, and biodiverse assemblage there, which is formed by both individual corals and their colonial relatives, together with accompanying biota (Roberts, Wheeler & Freiwald 2006; Stolarski et al. 2011). Such deep-water reefs grow slowly (over geological time scales), and in their skeletons’ annual bands have a record of shifts in the ocean’s climate; thus, they are significant paleo-environmental archives (Schröder-Ritzrau, Freiwald & Mangini 2005). Unfortunately, human direct activity, like bottom trawling, plastic pollution (both macro- and micro-) or the not fully understood implications of ocean acidification (e.g., for biomineralization), are highlighted as significant damage threats to these hidden ecosystems (Chapron et al. 2018; Doney et al. 2009; Roberts, Wheeler & Freiwald 2006).
Below the euphotic zone, there is insufficient light for photosynthesis in the twilight (dysphotic) zone, and below it, dark waters of the aphotic zone open, with no light penetration (see Fig. 1). Such a gradient of light in the water column affects not only the primary production or availability of tropical coral reef biodiversity hot spots, but also the biology – including adaptations for survival – of other organisms inhabiting aquatic environments. In the euphotic zone, similarly to the situation on land, organisms may rely on their sense of vision. This sense may be employed for communication, such as in mating or social behaviors, or displaying warnings: the blue-ringed octopus is famous for being one of the most venomous animals on Earth, and it quickly sends the warning by flashing the iridescent blue rings that appear on its body when disturbed (by muscle contraction mechanism; Mäthger et al. 2012). Also, for obtaining food, for example, when predators visually search for their prey, or hunt from concealment, or a supposed prey avoids becoming prey via camouflage, transparency, or migrations to dark areas of the water column (see review for camouflage–mimicry in fish by Randall 2005). Moonlight and artificial light (e.g., in areas such as ports or coastal towns) may impact the visibility and thus behavior of organisms that naturally hide in the darkness of night (e.g., Anokhina 2006; Burford & Robison 2020). Especially, the artificial light at night (ALAN), an expanding form of pollution, may impact a diverse group of organisms (attract/repel life forms and/or disrupt their biology). Some examples are: diel vertical migrations of organisms, coral symbiosis with dinoflagellates and spawning of these animals, behavior of hatchling turtles, behavior and interspecific interactions (like predation) of pelagic fish and krill, but also of sessile organisms in coastal areas (and even of seabirds) – all those are pivotal for survival of and condition of ecosystems (reviewed in Marangoni et al. 2022); (Bolton et al. 2017; Underwood, Davies & Queirós 2017).
Finally, the light in water may not only come from external, abiotic sources. The bioluminescence is an emission of visible light by a living being. A broad range of evolutionary lineages of organisms, from bacteria and protists, through cnidarians, crustaceans, mollusks, to the fish, have evolved the ability to produce light in marine environments (but is nearly absent in freshwaters). The classical example of bioluminescence used for luring prey in the darkness of the aphotic zone is the deep-water anglerfish – it has a long appendage on its head and in front of its mouth, finished with a glowing lure (thanks to symbiotic bacteria) that attracts prey (Haddock, Moline & Case 2010). However, the other example can be provided by one of the largest known deep-sea squid, Taningia danae, which was video recorded when hunted, at depths ranging from 240 m (at night) to 940 m (during the day). This squid appeared as an active, aggressive predator that was using the bioluminescence during the attack, which was interpreted as probably a blinding flash for the prey, and/or to measure target distance in its dark habitat (Kubodera, Koyama & Mori 2007). Another example is the Humboldt squid, an active and social high-level predator that spends its life in the dark waters below 200 meters. It uses bioluminescence, as presumed, for an advanced form of social communication. The visual signals of this squid enable it to rapidly convey and receive information between individuals, facilitating collective behaviors (Burford & Robison 2020).
Apart from light, the solar heating effects are mainly limited to the zone near the water surface.
Temperature gradient
Most of the solar heat is absorbed in the surface layers of water. The water layer, up to about 200 m deep in oceans and up to a dozen meters in lakes, known as the epipelagic zone, is also wind mixed. Temperatures there may vary more significantly according to the atmospheric impact (including climate and seasons) compared to the deeper parts of the water column (Kara, Rochford & Hurlburt 2003; Lampert & Sommer 1996).
Most of the biota that live in water are poikilothermic – “cold-blooded”, ectotherms (thus, except for some vertebrates, such as birds and mammals, which are homeothermic endotherms). Their body temperature is influenced by both the outside environment and the ambient temperature (Wagner et al. 2023). Water, with its high heat capacity and thermal conductivity, helps maintain the temperature of objects (e.g., bodies of aquatic organisms) close to the water temperature of their environment. The size of the organism can matter – smaller organisms may follow the temperature more easily than larger ones (Deutsch et al. 2022). Moreover, body temperature is a crucial factor in an organism’s physiology, affecting biological processes from biochemical reactions to metabolic efficiency, which can significantly influence survival. For example, the unicellular predatory protists (which are poikilothermic ectotherms) actively swim to hunt their prey. However, the temperature of their aquatic environment cannot be considered independently of its viscosity. Their swimming speed was experimentally proven to increase with temperature, and both the temperature-dependent physiology (metabolic rates) of these protists and the viscosity of the environment contributed to this result, which may have ecological and survival implications (Beveridge, Petchey & Humphries 2010). The other factors are the metabolism rate, O2 demand, and O2 availability. With body size increase, the organism needs to ensure that the surfaces of gas exchange are sufficiently large to compensate for its metabolic needs, and metabolism is a temperature-driven parameter (Deutsch et al. 2022). Unfortunately, the oxygen solubility declines with the growth of water temperature and with an increase in salt concentration (Green & Carritt 1967). Thus, the warmer the water is, the higher the metabolism rates of organisms become (and the metabolic O2 demand), and the more oxygen-limited the environment becomes. Thus, to compensate for these physical constraints, ectothermic organisms living in warmer conditions commonly mature and reproduce at smaller body sizes (and earlier) than in colder conditions (Forster, Hirst & Atkinson 2012), which can be a significant observation regarding global warming (Deutsch et al. 2022).
The water temperature gradient in a three-dimensional environment (warm surface and colder deeper waters) provides challenges for epipelagic fish physiology both daily and seasonally. Although the majority of ectotherms do not exhibit thermogenesis like endotherms, they can compensate for environmental temperature variations both behaviorally (by choosing the environment) and physiologically (by metabolism; Seebacher 2009). Nonetheless, some examples of larger fish have adaptations that enhance their ability to conserve metabolic heat (i.e., elevate tissue temperature above ambient), which may alter their possible depth of vertical diving and subsequent rewarming after feeding in cold waters. For example, a swordfish was observed to stay at depths for up to 8 hours a day, in temperatures 10 °C lower than those of the surface. Swordfish is known to have the anatomical structure – specialized tissue, a “brain heater” that warms up its brain and eye above the ambient temperature and protects the fish’s central neural system against rapid cooling when it dives deeply in colder waters (Carey 1982). At the same time, the blue sharks probably needed rewarming more often, because they oscillated back to the surface every few hours (more patterns of vertical movements of epipelagic fish were discussed in Andrzejaczek et al. 2019). On the other hand, the large size of the body helps stabilize temperature variations in its tissues as the large ectotherm animal moves through environments with diverse temperatures. The whale sharks – the largest fish – can dive into up to 1000 m depths, without decreasing their body temperature (thus without the metabolic costs) (Nakamura, Matsumoto & Sato 2020). Additionally, Bennett et al. (2021) consider that the thermal limits of organism survival are constrained by evolution and conserved through time across broad taxonomic groups, with a limited potential for further adaptation to increasing heat stress under current global warming.
Water density and pycnocline
The surface waters of the oceans (and lakes) are well mixed with wind as the dominant mechanism (but convection, evaporation, or tides may also be involved; Duxbury, Duxbury & Sverdrup 2002; Kara, Rochford & Hurlburt 2003). The depth of this layer varies seasonally and by climatic zone. This is called a Mixed Layer Depth, which is important transitional layer – acting as boundary between deeper waters and the atmosphere above, regulating temperature changes or availability of nutrients; Abdulla et al. 2018; Kara, Rochford & Hurlburt 2003). The waters below 1000 m in the ocean are uniformly cold, dense, and salty. Between mixed surface waters and those homogeneous in parameters, we observe a water layer where density (defined by temperature and salinity gradients) increases rapidly, acting as a physical barrier to vertical mixing of seawater. This layer is called the pycnocline. Pycnocline is commonly linked to thermocline (rapid decrease of temperature up to about 0-4 oC, which is a standard temperature of deep waters; Emery & Meincke 1986) and halocline (rapid increase of salinity) – because the density of water is related to temperature, and to its salinity (Fiedler et al. 2013). The cold waters sink and stay deep because they are denser than the warmer waters. The warm waters of the surface have limited nutrient availability (almost all nutrients are already consumed by phytoplankton and other participants of food webs), and the pycnocline does not allow the nutrients that are dissolved in deep waters to migrate to the surface (Bristow et al. 2017). However, the nutrient-rich deep waters may flux through the pycnocline toward the surface, leading to a decrease in surface temperature as a side effect, but primarily to an important nutrient supply for phytoplankton – a base for the food web.
The movement of deep waters upward is generally possible along a coastline11 when strong winds force the surface waters to move toward the ocean, allowing the released space to be filled by the waters pulled from the deep. This process, called upwelling, wherever it occurs, ensures high biological productivity in the ocean and related environments, including the production of exploitable fish important for human food and trade (Bakun et al. 2010; NOAA 2024d; Ryther 1969). The current climate crisis can affect these water transfers. Xiu et al. (2018) explain the decline trend of the average biomass and productivity of phytoplankton as a possible result of warming of the surface waters. The heated surface waters are harder to mix with deeper layers, enhancing water stratification and limiting the flux of nutrients. However, the results from the California Current System showed increased upwelling intensity due to strong winds in the coastal region, along with stronger stratification. Alternatively, warmer water may increase the metabolic rate of ectotherm animals that feed on phytoplankton, thereby shifting the food web structure (O’Connor et al. 2009). The anthropogenic impact can, potentially, touch the natural cycles even more directly. The offshore wind farms have, of late, been recognized as a valuable source of “green energy”. Raghukumar et al. (2023) recently asked a question about the possible relation between offshore wind farm activity and the winds that drive local upwelling. They modelled the changes in the wind currents by wind turbines, on the California coast. Although they reported that the model suggested turbines promoted a wind stress curl-driven upwelling, the impact on the coastal region was minimal. Nevertheless, regarding the essential influence of nutrient fluxes on fishery and ecosystem structure, which are currently unknown for the cited research, the authors underline the importance of expanding environmental analyses before placing any anthropogenic structures that may interact with natural cycles.
3. Human activities and ocean risks
Some examples of the human-environment interactions were already described in the sections above. However, this place provides a summary of the most challenging factors affecting life under the sea surface.
Global warming
Global warming is the phenomenon of observation of an unusually rapid rise in Earth’s average surface temperature (observed during at least a few-decade sample – a period long enough to remove the interannual variations), and when compared to a reference period (the IPCC (2021) indicates a baseline for 1850-1900). Such a rapid rise, faster than ever in the instrumental record, reached the temperature increase of 0.27 °C per decade in 2015–2024 (Forster et al. 2025). This, despite discussions about the degree of the natural input to this phenomenon (see analysis in Keller 2003), is widely agreed to be a result of the increased release (thus increased concentration in the atmosphere) of the greenhouse gases, including CO2, by human activity since the industrial era – e.g., burning the fossil fuels (Al-Ghussain 2019; Houghton 2001; Kerr 2007). The currently observed global warming may lead to and be a part of the change of global or regional climate patterns – a climate change (those two terms are commonly used informally as synonyms, while they describe slightly different phenomena; IPCC 2021; Lineman et al. 2015). Some effects of the rise in temperatures include the warming of the ocean and the melting of ice and glaciers worldwide, which lead to a rise in sea level, increased evaporation, and ocean acidification.
Forster et al. (2025) summarizes that the temperature anomaly in oceans, according to reference years, was estimated to 1.02 oC in 2015-2024, and that the most impacted layer of the oceans is the surface layer (up to 700 m), then the heat propagates to deeper waters (700-2000 m) with the most negligible impact on deep waters below 2000 m. The warmer water is altering the ectotherm organisms’ metabolism, including oxygen demand (and less oxygen is available in warm water). This may induce mobility in organisms in search of available habitats, both as an escape from unfavorable conditions and as an expansion into new areas (previously not physiologically available) – which may change the ranges of aquatic organisms and reshape local and global ecosystems (also by negative impacts e.g. increased competition or spreading disease), and even lead to the socioeconomic consequences (Strang & Bosker 2024). Eventually, if the extreme heat event waves appear, accompanying the global warming, they may lead to indirect or direct influence on aquatic ecosystems, e.g., long-lasting blooms of mucilaginous algae (that may cover and disturb other organisms), or directly weaken or kill aquatic organisms, especially benthic sessile animals from shallow waters, like corals (Al-Ghussain 2019; Lough, Anderson & Hughes 2018; von Schuckmann et al. 2019). Finally, warmer oceanic water accelerates evaporation, influencing the global water cycle and greenhouse effect (making the atmosphere more humid) and potentially leading to more extreme weather events (Al-Ghussain 2019).
The predicted rise of the sea level would be approximately half a meter before the end of the 21st century (without reducing the climate crisis actions; Houghton 2001). This will affect the coastal ecosystems (e.g., by local changes in nutrient availability or increased sedimentation) and will also alter the range of tides or the destructive effects of storms (Al-Ghussain 2019; Noe et al. 2013). During the period from 2019 to 2024, the sea level rose by an average of 2.5 cm; although it was rising during the 20th century, the current rise is more rapid (Forster et al. 2025). The ocean water is absorbing the carbon dioxide – CO2 (this, in fact, reduces the extent of global warming). When it happens, the carbonic acid (H2CO3) is generated, ions of which act as the ocean’s buffer. However, when more CO2 is available, more ions of H+ (“acidic particles”) are formed, thus pH of the water drops. The current value of the mean oceanic pH is 8.1, which is 0.1 lower than the reference from the late 18th century (NOAA 2025b). This may lead to disturbances in the physiological processes of organisms and negatively influence biomineralization (such as in corals and other living beings that produce a skeleton). In the broader context, this may reshape community assemblages in different marine ecosystems (Logan 2010).
Marine pollution
The progress of urbanization – especially of the coastlines – led to human–ocean interaction, resulting in commonly rebuilt, reduced, and polluted areas. The pollution consists of the traditionally understood introduction of waste objects and liquids into the marine ecosystem, as well as pollution from noise, light, or interactions. Underwater noise from human activities, mainly from boats, is present worldwide. During the COVID-19 pandemic, human activity at sea decreased (called the “Anthropause”), causing a significant reduction in ocean human-made noise. This altered the behavioral activity, echolocation responses, whistle repertoire, and attention of dolphins in Bermuda (Stevens, Allen & Bruck 2023), and Panama (Gagne et al. 2022). It also increased (by double) the presence of monk seals in Hawai’i and evidenced the inhibitory effect of humans (when active) on reef ecosystems there (Madin et al. 2025).
The source of pollution may originate from terrestrial environments (about 80% of waste!), the sea itself, or the air. Ma et al. (2023) explains that the air pollution commonly starts with picking up the tiny particles from the land, but gases and chemical substances are also part of the air pollution category. Sea sources of pollution are primarily the effects of the fishery industry. Finally, land source can come from a broad area or as a point on a coastline. This is an effect of a wide variety of human activities, from agriculture and industry to households and human behaviors and habits. In this category fall the input of nutrients, which results in, for example, the eutrophication of coral reefs in urbanized areas, leading to a decrease in coral cover and the development of algal blooms and other potentially harmful organisms, such as certain invertebrates (Duprey, Yasuhara & Baker 2016; Girard et al. 2025). Plastic pollution is the other challenge to modern ecosystems. Plastics are a broad group of artificial polymers, widely used since the middle of 20th century. The biological complication with these materials is that they are not easily decomposed, so after being introduced to the environment, they may remain there for decades or centuries (e.g. EPA 2025). Plastics may become consumed and be introduced to the food web (but particles may harm intestines or mimic the stomach being full, causing the organisms to die from starvation) or provide a surface for settlement, both for macro and microorganisms. Microplastics (produced in sizes below 5 mm, or originating from fragmentation) and plastic debris are regarded as an emerging and severe anthropogenic threat to the aquatic ecosystems worldwide (Sharma, Sharma & Chatterjee 2021; Wright, Thompson & Galloway 2013). Nevertheless, new plastic objects in the ecosystem may be employed to serve ecological functions. The hermit crabs can use them as a shelter, in place of the gastropod shell (Jagiello, Dylewski & Szulkin 2024), and similar behavior was observed in octopus (however, they preferred glass objects a little more; Freitas et al. 2022). The sea urchin (Lytechinus variegatus) collects plastic litter for (likely) protection, camouflage, and ballast (Barros et al. 2020).
Unsustainable fishing
The seas and oceans are the primary source of fish for the food industry. However, harvesting fish populations to the extent that they cannot replenish, known as overfishing, leads to significant disturbances in their condition and a decline in their biomass. Overfished populations may be prone to disease due to elevated stress level, which can additionally ease their decline. Overfishing is affecting biodiversity and may also pose a risk to food security and the lives of those relying on this resource – if fish become too rare (Wright 2023). The North Pacific, commercially important, blue tuna is one of the species whose population suffered from decades of fishing, leading to low stock for years (Nakatsuka et al. 2017).
The over-exploitation of cod in the Baltic Sea12 led to a critical decline in the populations of this fish. Birgersson, Söderström & Belhaj (2022) recommend an ecosystem-based approach to aid the recovery of this species, including monitoring and protection of habitats for all life stages on one hand, and low-impact fisheries on the other hand. Separation of the cultural, economic, and social, as well as biological aspects of the sustainability of fisheries is difficult, but broader attempts may aid in the recovery of over-exploited fish populations worldwide.
Summary
Life evolved on our planet thanks to the availability of water. Water plays a central role in climate regulation and the hydrological cycle, and it creates diverse environments and habitats – all of which depend on its physical characteristics. These include the gradients of parameters like light, temperature, density, or salinity. Representatives of the majority of the organism’s lineages live in water and have evolved adaptations for the aquatic habitat. This may appear challenging to realize from the perspective of a terrestrial organism; however, advances in research have provided a better understanding of how underwater life functions, interacts with other biota and with its environment, and ultimately survives in the face of human activity.
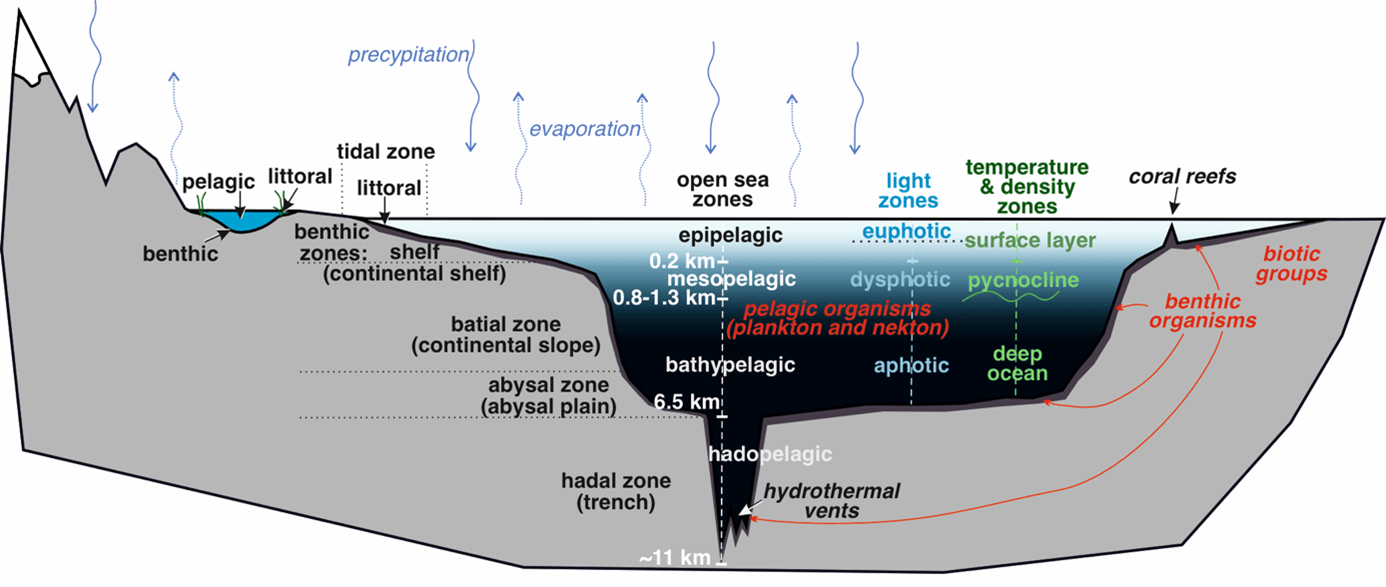
Figure 1. The water on Earth in oceans (and lakes) – vertical water divisions, life zones, and biotic groups, with the water cycle (simplified); information summarized, based on (Cox, Moore & Ladle 2016; Duxbury, Duxbury & Sverdrup 2002; Meerhoff & González-Sagrario 2021; NOAA 2024b; Oki & Kanae 2006; Weiner 1999).
Literature
Abdulla, C. P., Alsaafani, M. A., Alraddadi, T. M. & Albarakati, A. M., 2018. ‘Mixed layer depth variability in the Red Sea’. Ocean Science, 14, 4: 563-573.
Abe, Y., 1993. ‘Physical state of the very early Earth. Lithos, 30, 3-4: 223-235.
Abe, Y. & Matsui, T., 1985. ‘The formation of an impact-generated H2O atmosphere and its implications for the early thermal history of the Earth’. Journal of Geophysical Research: Solid Earth, 90, S02: C545-C559.
Al-Ghussain, L., 2019. ‘Global warming: review on driving forces and mitigation’. Environmental Progress & Sustainable Energy, 38, 1: 13-21.
Allègre, C. J., Manhès, G. & Göpel, C., 1995. ‘The age of the Earth’. Geochimica et Cosmochimica Acta, 59, 8: 1445-1456.
Andrzejaczek, S., Gleiss, A. C., Pattiaratchi, C. B. & Meekan, M. G., 2019. ‘Patterns and drivers of vertical movements of the large fishes of the epipelagic’. Reviews in Fish Biology and Fisheries, 29, 2: 335-354.
Anokhina, L. L., 2006. ‘Influence of moonlight on the vertical migrations of benthopelagic organisms in the near-shore area of the Black Sea’. Oceanology, 46, 3: 385-395.
Arabeyyat, Z. H., Sweiss, M. A., Alsoqi, F. M., Kteifan, M., Al-Horani, F. & Taha, Z. D., 2024. ‘Molecular identification of dinoflagellates symbionts (Zooxanthellae) in corals of the Jordanian Gulf of Aqaba, Red Sea’. Regional Studies in Marine Science, 77, 103649.
Babcock, R. C., Wills, B. L. Simpson, C. J., 1994. ‘Mass spawning of corals on a high latitude coral reef’. Coral Reefs, 13, 3: 161-169.
Baker, L. B., 2019. ‘Physiology of sweat gland function: the roles of sweating and sweat composition in human health’. Temperature, 6, 3: 211-259.
Bakun, A., Field, D. B., Redondo-Rodriguez, A. & Weeks, S. J., 2010. ‘Greenhouse gas, upwelling-favorable winds, and the future of coastal ocean upwelling ecosystems’. Global Change Biology, 16, 4: 1213-1228.
Barber, J. & Tran, P. D., 2013. ‘From natural to artificial photosynthesis’. Journal of the Royal Society Interface, 10, 81: 20120984.
Barber, R. & Hilting, A., 2000. ‘Achievements in biological oceanography’. In: National Research Council (US) Ocean Studies Board. (eds.) . 50 Years of Ocean Discovery: National Science Foundation 1950—2000. Washington (DC): National Academies Press (US), pp. 11-21.
Barros, F., Santos, D., Reis, A., Martins, A., Dodonov, P. & Nunes, J., 2020. ‘Choosing trash instead of nature: Sea urchin covering behavior’. Marine Pollution Bulletin, 155, 111188.
Bennett, J. M. Sunday, J., Calosi, P., Villalobos, F., Martínez, B., Molina-Venegas, R., Araújo, M. B., Algar, A. C., Clusella-Trullas, S., Hawkins, B. A., Keith, S. A., Kühn, I., Rahbek, C., Rodríguez, L., Singer, A., Morales-Castilla, I. & Olalla-Tárraga, M. Á., 2021. ‘The evolution of critical thermal limits of life on Earth’. Nature Communications 12, 1: 1198.
Beveridge, O. S., Petchey, O. L. & Humphries, S., 2010. ‘Mechanisms of temperature-dependent swimming: the importance of physics, physiology and body size in determining protist swimming speed’. Journal of Experimental Biology, 213, 24: 4223-4231.
Birgersson, L.,Söderström, S. & Belhaj, M., 2022. The decline of cod in the Baltic Sea – a review of biology, fisheries and management, including recommendations for cod recovery. Stockholm, Sweden: The Fisheries Secretariat.
Bolton, D., Mayer-Pinto, M., Clark, G. F., Dafforn, K. A., Brassil, W. A., Becker, A. & Johnston, E. L., 2017. ‘Coastal urban lighting has ecological consequences for multiple trophic levels under the sea’. Science of The Total Environment, 576, 1-9.
Boström-Einarsson, L., Babcock, R. C., Bayraktarov, E., Ceccarelli, D., Cook, N., Ferse, S. C. A., Hancock, B., Harrison, P., Hein, M., Shaver, E., Smith, A., Suggett, D., Stewart-Sinclair, P. J., Vardi, T. & McLeod, I. M., 2020. ‘Coral restoration – a systematic review of current methods, successes, failures and future directions’. PLoS One, 15, 1: e0226631.
Brandl, S. J., Rasher, D. B., Côté, I. M., Casey, J. M., Darling, E. S., Lefcheck, J. S. & Duffy, J. E., 2019. ‘Coral reef ecosystem functioning: eight core processes and the role of biodiversity’. Frontiers in Ecology and the Environment, 17, 8: 445-454.
Brandt, M. E. & McManus, J. W., 2009. ‘Disease incidence is related to bleaching extent in reef-building corals’. Ecology, 90, 10: 2859-2867.
Bristow, L. A., Mohr, W.,Ahmerkamp, S. & Kuypers, M. M. M., 2017. ‘Nutrients that limit growth in the ocean’. Current Biology, 27, 11: R474-R478.
Bruckner, A. W., 2002. ‘Life-saving products from coral reefs’. Issues in Science and Technology, 18, 3:39-44.
Brusca, R., Moore, W. & Shuster, S., 2016. Invertebrates. 3rd ed. Oxford University Press.
Burford, B. P. & Robison, B. H., 2020. ‘Bioluminescent backlighting illuminates the complex visual signals of a social squid in the deep sea’. Proceedings of the National Academy of Sciences, 117, 15: 8524-8531.
Burke, L., Reytar, K. Spalding, M. & Perry, A., 2012. Reefs at risk revisited in the coral triangle. World Resources Institute.
Carey, F. G., 1982. ‘A brain heater in the swordfish’. Science, 216, 4552: 1327-1329.
Carmignani, A., Skrzypek, G., Brooker, R. M., Meekan, M. G., Chase, T. J., Shantz, A. A. & Barneche, D. R., 2025. ‘The relationship between nutrient supply from resident fishes and the growth, condition, and thermal tolerance of corals’. Coral Reefs, 44, 5: 1815-183.
Chaplin, M., 2006. ‘Do we underestimate the importance of water in cell biology?’. Nature Reviews Molecular Cell Biology 7, 11: 861-6.
Chapron, L., Peru, E., Engler, A., Ghiglione, J. F., Meistertzheim, A. L., Pruski, A. M., Purser, A., Vétion, G., Galand, P. E. & Lartaud, F., 2018. ‘Macro- and microplastics affect cold-water corals growth, feeding and behaviour’. Scientific Reports, 8, 1: 15299.
Corrochano-Fraile, A., Carboni, S., Green, D. M., Taggart, J. B., Adams, T. P., Aleynik, D. & Bekaert, M., 2024. ‘Estimating blue mussel (Mytilus edulis) connectivity and settlement capacity in mid-latitude fjord regions’. Communications Biology, 7, 1: 166.
Costello, M. J. & Chaudhary, C., 2017. ‘Marine biodiversity, biogeography, deep-sea gradients, and conservation’. Current Biology, 27, 11: R511-R527.
Cox, C. B., Moore, P. D. & Ladle, R. J., 2016. Biogeography. An ecological and evolutionary approach. Chichester, UK; Hoboken, NJ: John Wiley & Sons.
Deutsch, C., Penn, J. L., Verberk, W. C. E. P., Inomura, K., Endress, M.-G. & Payne, J. L., 2022. ‘Impact of warming on aquatic body sizes explained by metabolic scaling from microbes to macrofauna’. Proceedings of the National Academy of Sciences, 119, 28: e2201345119.
Dodd, M. S., Papineau, D., Grenne, T., Slack, J. F., Rittner, M., Pirajno, F., O’Neil, J. & Little, C. T. S., 2017. ‘Evidence for early life in Earth’s oldest hydrothermal vent precipitates’. Nature, 543, 7643: 60-64.
Doney, S. C., Fabry, V. J., Feely, R. A. & Kleypas, J. A., 2009. ‘Ocean acidification: the other CO2 problem’. Annual Review of Marine Science, 1, 169-192.
Donohue, C. G., Hemmi, J. M. & Kelley, J. L., 2020. ‘Countershading enhances camouflage by reducing prey contrast’. Proceedings of the Royal Society B: Biological Sciences, 287, 1927: 20200477.
Drake, M. J., 2005. ‘Origin of water in the terrestrial planets’. Meteoritics & Planetary Science, 40, 4: 519-527.
Duprey, N. N., Yasuhara, M. & Baker, D. M., 2016. ‘Reefs of tomorrow: eutrophication reduces coral biodiversity in an urbanized seascape’. Global Change Biology, 22, 11: 3550-3565.
Duxbury, A. C., Duxbury, A. B. & Sverdrup, K. A., 2002. Oceany Świata. Warszawa: Wydawnictwo Naukowe PWN.
Edmondson, W. T., 1956. ‘The relation of photosynthesis by phytoplankton to light in lakes’. Ecology, 37, 1: 161-174.
Elster, J. & Benson, E., 2004. ‘Life in the polar terrestrial environment with a focus on algae and cyanobacteria’. In: B. J. Fuller, N. Lane and Benson E. (eds), Life in the frozen state. Boca Raton: CRC Press pp. 111-150.
Emery, W. J. & Meincke, J. P., 1986. ‘Global water masses: summary and review’. Oceanologica Acta, 9, 4: 383-391.
EPA, 2025. Impacts of Plastic Pollution. US Environmental Protection Agency Webpage, viewed: 28 September 2025, <https://www.epa.gov/plastics/impacts-plastic-pollution>.
Fiedler, P. C., Mendelssohn, R., Palacios, D. M. & Bograd, S. J., 2013. ‘Pycnocline variations in the eastern tropical and North Pacific, 1958–2008’. Journal of Climate, 26, 2: 583-599.
Forster, J., Hirst, A. G. & Atkinson, D., 2012. ‘Warming-induced reductions in body size are greater in aquatic than terrestrial species’. Proceedings of the National Academy of Sciences, 109, 47: 19310-19314.
Forster, P. M., Smith, C., Walsh, T., Lamb, W. F., Lamboll, R., Cassou, C., Hauser, M., Hausfather, Z., Lee, J. Y., Palmer, M. D., von Schuckmann, K., Slangen, A. B. A., Szopa, S., Trewin, B., Yun, J., Gillett, N. P., Jenkins, S., Matthews, H. D., Raghavan, K., Ribes, A., Rogelj, J., Rosen, D., Zhang, X., Allen, M., Aleluia Reis, L., Andrew, R. M., Betts, R. A., Borger, A., Broersma, J. A., Burgess, S. N., Cheng, L., Friedlingstein, P., Domingues, C. M., Gambarini, M., Gasser, T., Gütschow, J., Ishii, M., Kadow, C., Kennedy, J., Killick, R. E., Krummel, P. B., Liné, A., Monselesan, D. P., Morice, C., Mühle, J., Naik, V., Peters, G. P., Pirani, A., Pongratz, J., Minx, J. C., Rigby, M., Rohde, R., Savita, A., Seneviratne, S. I., Thorne, P., Wells, C., Western, L. M., van der Werf, G. R., Wijffels, S. E., Masson-Delmotte, V. & Zhai, P., 2025. ‘Indicators of global climate change 2024: annual update of key indicators of the state of the climate system and human influence’. Earth System Science Data, 17, 6: 2641-2680.
Frankowiak, K., Wang, X. T., Sigman, D. M., Gothmann, A. M., Kitahara, M. V., Mazur, M., Meibom, A. & Stolarski, J., 2016. ‘Photosymbiosis and the expansion of shallow-water corals’. Science Advances, 2, 11:e1601122.
Freitas, T. B. N., Leite, T. S., de Ramos, B., di Cosmo, A. & Proietti, M. C. 2022. ‘In an octopus’s garden in the shade: underwater image analysis of litter use by benthic octopuses’. Marine Pollution Bulletin, 175, 113339.
Gagne, E., Perez-Ortega, B., Hendry, A. P., Melo-Santos, G., Walmsley, S. F., Rege-Colt, M., Austin, M. & May-Collado, L. J., 2022. ‘Dolphin communication during widespread systematic noise reduction-a natural experiment amid COVID-19 lockdowns’. Frontiers in Remote Sensing, 3, 934608.
GBRF, n.d.. Coral Bleaching. Great Barrier Reef Foundation Website, viewed: 28 September 2025, <https://www.barrierreef.org/the-reef/threats/coral-bleaching>.
Genda, H. & Ikoma, M., 2008. ‘Origin of the ocean on the Earth: early evolution of water D/H in a hydrogen-rich atmosphere’. Icarus, 194, 1: 42-52.
Girard, E. B., Pratama, A. M. A., del Rio-Hortega, L., Volkenandt, S., Macher, J.-N. & Renema, W., 2025. ‘Coastal eutrophication transforms shallow micro-benthic reef communities’. Science of The Total Environment, 961, 178252.
Green, E. J. & Carritt, D. E., 1967. ‘Oxygen solubility in sea water: thermodynamic influence of sea salt’. Science, 157, 3785: 191-193.
Grosberg, R. K., Vermeij, G. J. & Wainwright, P. C., 2012. ‘Biodiversity in water and on land’. Current Biology, 22, 21: R900-R903.
Grott, M., Morschhauser, A., Breuer, D. & Hauber, E., 2011. ‘Volcanic outgassing of CO2 and H2O on Mars’. Earth and Planetary Science Letters, 308, 3: 391-400.
Haddock, S. H. D., Moline, M. A. & Case, J. F., 2010. ‘Bioluminescence in the sea’. Annual Review of Marine Science, 2, 443-493.
Hata, T., Madin, J. S., Cumbo, V. R., Denny, M., Figueiredo, J., Harii, S., Thomas, C. J. & Baird, A. H., 2017. ‘Coral larvae are poor swimmers and require fine-scale reef structure to settle’. Scientific Reports, 7, 1: 2249.
Houghton, J., 2001. ‘The science of global warming’. Interdisciplinary Science Reviews, 26, 4: 247-257.
Hughes, T. P., Barnes, M. L., Bellwood, D. R., Cinner, J. E., Cumming, G. S., Jackson, J. B. C., Kleypas, J., van de Leemput, I. A., Lough, J. M., Morrison, T. H., Palumbi, S. R., van Nes, E. H. & Scheffer, M., 2017. ‘Coral reefs in the Anthropocene’. Nature, 546, 7656: 82-90.
Hwang, H.-s., Cho, B., Cho, J., Park, B. & Kim, T., 2022. ‘New record of hydrothermal vent squat lobster (Munidopsis lauensis) provides evidence of a dispersal corridor between the Pacific and Indian Oceans’. Journal of Marine Science and Engineering, 10, 3: 400.
IPCC, 2021. Annex VII: Glossary. In: J. B. R. Matthews, V. Möller, R. van Diemen, J.S, Fuglestvedt, V. Masson-Delmotte, C. Méndez, S. Semenov & Reisinger, A. (eds), Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change [Masson-Delmotte, V., P. Zhai, A. Pirani, S.L. Connors, C. Péan, S. Berger, N. Caud, Y. Chen, L. Goldfarb, M.I. Gomis, M. Huang, K. Leitzell, E. Lonnoy, J.B.R. Matthews, T.K. Maycock, T. Waterfield, O. Yelekçi, R. Yu, & Zhou, B. (eds.)]. Cambridge, UK and New York, USA: Cambridge University Press, pp. 2215–2256.
Jagiello, Z., Dylewski, Ł. & Szulkin, M., 2024. ‘The plastic homes of hermit crabs in the Anthropocene’. Science of The Total Environment, 913, 168959.
Kadlec, R. H., 2006. ‘Water temperature and evapotranspiration in surface flow wetlands in hot arid climate’. Ecological Engineering, 26, 4: 328-340.
Kara, A. B., Rochford, P. A. & Hurlburt, H. E., 2003. ‘Mixed layer depth variability over the global ocean’. Journal of Geophysical Research: Oceans, 108, C3: 3079.
Kasting, J. F., Whitmire, D. P. & Reynolds, R. T., 1993. ‘Habitable zones around main sequence stars’. Icarus, 101, 1: 108-128.
Keller, C. F., 2003. ‘Global warming: the balance of evidence and its policy implications. a review of the current state-of-the-controversy’. Scientific World Journal, 3, 357-411.
Kerr, R. A., 2007. ‘Global warming is changing the world’. Science, 316, 5822: 188-190.
Kleypas, J. A., McManus, J. W. & Menez, L. A. B., 1999. ‘Environmental limits to coral reef development: where do we draw the line?’ American Zoologist, 39, 1: 146-159.
Kontogeorgis, G. M., Holster, A., Kottaki, N., Tsochantaris, E., Topsøe, F., Poulsen, J., Bache, M., Liang, X., Blom, N. S. & Kronholm, J., 2022. ‘Water structure, properties and some applications – a review’. Chemical Thermodynamics and Thermal Analysis, 6, 100053.
Kubodera, T., Koyama, Y. & Mori, K., 2007. ‘Observations of wild hunting behaviour and bioluminescence of a large deep-sea, eight-armed squid, Taningia danae’. Proceedings of the Royal Society of London. Series B, Biological Sciences, 274, 1613: 1029-34.
Lampert, W. & Sommer, U., 1996. Ekologia wód śródlądowych. Warszawa: Wydawnictwo Naukowe PWN.
Lee, Z., Weidemann, A., Kindle, J., Arnone, R., Carder, K. L. & Davis, C., 2007. ‘Euphotic zone depth: its derivation and implication to ocean-color remote sensing’. Journal of Geophysical Research: Oceans, 112, C3: C03009.
Leite Jardim, V., Boyé, A., Le Garrec, V., Maguer, M., Tauran, A., Gauthier, O. & Grall, J. (2025). ‘Habitat complexity promotes species richness and community stability: a case study in a marine biogenic habitat’. Oikos, 2025, 7: e10675.
Lineman, M., Do, Y., Kim, J. Y. & Joo, G.-J., 2015. ‘Talking about climate change and global warming’. PLoS One, 10, 9: e0138996.
Liu, J. & van Iersel, M. W., 2021. ‘Photosynthetic physiology of blue, green, and red light: light intensity effects and underlying mechanisms’. Frontiers in Plant Science 12, 619987.
Logan, C. A., 2010. ‘A review of ocean acidification and america’s response’. BioScience, 60, 10: 819-828.
Lörz, A. N., Schwentner, M., Bober, S. & Jażdżewska, A. M., 2023. ‘Multi-ocean distribution of a brooding predator in the abyssal benthos’. Scientific Reports, 13, 1: 15867.
Lough, J. M., Anderson, K. D. & Hughes, T. P. 2018. ‘Increasing thermal stress for tropical coral reefs: 1871–2017’. Scientific Reports, 8, 1: 6079.
Lutzenkirchen, L. L., Duce, S. J. & Bellwood, D. R., 2024. ‘Exploring benthic habitat assessments on coral reefs: a comparison of direct field measurements versus remote sensing’. Coral Reefs, 43, 2: 265-280.
Ma, J., Ma, R., Pan, Q., Liang, X., Wang, J. & Ni, X., 2023. ‘A global review of progress in remote sensing and monitoring of marine pollution’. Water, 15, 19: 3491.
Madin, E. M. P., Suan, A., Severino, S. J. L., Tsang, A. O., Madin, J. S., Rago, Z., Gerber, I. & Rodgers, K. S., 2025. ‘COVID-19 anthropause affects coral reef ecosystems through biophysical changes’. npj Ocean Sustainability., 4, 1: 48.
Mamajek, E. E., Prsa, A., Torres, G., Harmanec, P., Asplund, M., Bennett, P. D., Capitaine, N., Christensen-Dalsgaard, J., Depagne, E., Folkner, W. M., Haberreiter, M., Hekker, S., Hilton, J. L., Kostov, V., Kurtz, D. W., Laskar, J., Mason, B. D., Milone, E. F., Montgomery, M. M., Richards, M. T., Schou, J. & Stewart, S. G., 2015. ‘IAU 2015 Resolution B3 on Recommended Nominal Conversion Constants for Selected Solar and Planetary Properties’. arXiv, arXiv:1510.07674v1 [astro-ph.SR].
Maragos, J. E., Crosby, M. P. & McManus, J. W., 1996. Coral reefs and biodiversity: a critical and threatened relationship’. Oceanography, 9, 1: 83-99.
Marangoni, L. F. B., Davies, T., Smyth, T., Rodríguez, A., Hamann, M., Duarte, C., Pendoley, K., Berge, J., Maggi, E. & Levy, O., 2022. Impacts of artificial light at night in marine ecosystems—a review. Global Change Biology, 28, 18: 5346-5367.
Marra, J. F., Lance, V. P., Vaillancourt, R. D. & Hargreaves, B. R., 2014. ‘Resolving the ocean’s euphotic zone’. Deep Sea Research Part I: Oceanographic Research Papers, 83, 45-50.
Marty, B., 2012. ‘The origins and concentrations of water, carbon, nitrogen and noble gases on Earth’. Earth and Planetary Science Letters, 313-314, 56-66.
Mäthger, L. M., Bell, G. R. R., Kuzirian, A. M., Allen, J. J. & Hanlon, R. T., 2012. ‘How does the blue-ringed octopus (Hapalochlaena lunulata) flash its blue rings?’ Journal of Experimental Biology, 215, 21: 3752-3757.
McManus, M. A. & Woodson, C. B., 2012. ‘Plankton distribution and ocean dispersal’. Journal of Experimental Biology, 215, 6: 1008-1016.
Meech, K. & Raymond, S., 2019. ‘Origin of Earth’s water: sources and constraints’. arXiv, arXiv:1912.04361v1.
Meerhoff, M. & González-Sagrario, M., 2021. ‘Habitat complexity in shallow lakes and ponds: importance, threats, and potential for restoration’. Hydrobiologia, 849, 3737–3760.
Meesters, E. H. & Bak, R. P. M., 1993. ‘Effects of coral bleaching on tissue regeneration potential and colony survival’. Marine Ecology Progress Series, 96, 2: 189-198.
Meran, G., Siehlow, M. & von Hirschhausen, C., 2021. ‘Water availability: a hydrological view’. In: G. Meran, M. Siehlow & C. von Hirschhausen (eds). The economics of water: rules and institutions. Cham: Springer International Publishing, pp. 9-21.
Mojzsis, S. J., Harrison, T. M. & Pidgeon, R. T., 2001. ‘Oxygen-isotope evidence from ancient zircons for liquid water at the Earth’s surface 4,300 Myr ago’. Nature, 409, 6817: 178-81.
Muldrew, K., Acker, J., Elliot, J. A. W. & McGann, L., 2004. ‘The water to ice transition: implications for living cells’. In: B. J. Fuller, N. Lane & Benson, E. (eds), Life in the Frozen State. Boca Raton: CRC Press, pp. 67-108.
Nakamura, I., Matsumoto, R. Sato, K., 2020. ‘Body temperature stability in the whale shark, the world’s largest fish’. Journal of Experimental Biology, 223, 11: jeb210286.
Nakatsuka, S., Ishida, Y., Fukuda, H. & Akita, T., 2017. ‘A limit reference point to prevent recruitment overfishing of pacific bluefin tuna’. Marine Policy, 78, 107-113.
NASA, 2025. Earth. NASA, viewed: 28 September 2025, <https://science.nasa.gov/earth/facts/>.
NOAA, 2024a. How Deep Is the Ocean?. National Ocean Service Website, viewed: 28 September 2025, <https://oceanservice.noaa.gov/facts/oceandepth.html>.
NOAA, 2024b. How Far Does Light Travel in the Ocean?. National Ocean Service Website, viewed: 28 September 2025, <https://oceanservice.noaa.gov/facts/light_travel.html>.
NOAA, 2024c. In What Types of Water Do Corals Live?. National Ocean Service Website, viewed: 28 September 2025, , < https://oceanservice.noaa.gov/facts/coralwaters.html>.
NOAA, 2024d. What Is Upwelling?. National Ocean Service Website, viewed: 28 September 2025, < https://oceanservice.noaa.gov/facts/upwelling.html>.
NOAA, 2024e. Why Are Coral Reefs Important? National Ocean Service Website, viewed: 28 September 2025, <https://oceanservice.noaa.gov/education/tutorial_corals/coral07_importance.html>.
NOAA, 2025a. How Much of the Ocean Has Been Explored?. National Ocean Service Website, viewed: 28 September 2025, <https://oceanexplorer.noaa.gov/ocean-fact/explored/> .
NOAA, 2025b. Ocean Acidification. National Ocean Service Website, viewed: 1 October 2025, <https://www.noaa.gov/education/resource-collections/ocean-coasts/ocean-acidification>.
Noe, G. B., Krauss, K. W., Lockaby, B. G., Conner, W. H. & Hupp, C. R., 2013. ‘The effect of increasing salinity and forest mortality on soil nitrogen and phosphorus mineralization in tidal freshwater forested wetlands’. Biogeochemistry, 114, 1: 225-244.
Nutman, A. P., Bennett, V. C., Friend, C. R., Van Kranendonk, M. J. & Chivas, A. R., 2016. ‘Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures’. Nature, 537, 7621: 535-538.
O’Connor, M. I., Piehler, M. F., Leech, D. M., Anton, A. & Bruno, J. F., 2009. ‘Warming and resource availability shift food web structure and metabolism’. PLoS Biology, 7, 8: e1000178.
Oki, T. & Kanae, S., 2006. ‘Global hydrological cycles and world water resources’. Science, 313, 5790: 1068-1072.
Paps, J., Rossi, M. E., Bowles, A. M. C. & Álvarez-Presas, M., 2023. ‘Assembling animals: trees, genomes, cells, and contrast to plants’. Frontiers in Ecology and Evolution, 11, 1185566.
Perlman, H. & Evans, J., 2019. The natural water cycle. USGS, viewed: 26 August 2025, <https://www.usgs.gov/media/images/natural-water-cycle-jpg>.
Piani, L., Marrocchi, Y., Rigaudier, T., Vacher, L. G., Thomassin, D. & Marty, B., 2020. ‘Earth’s water may have been inherited from material similar to enstatite chondrite meteorites. Science, 369, 6507: 1110-1113.
Pineda, J., Hare, J. A. & Sponaugle, S. U., 2007. ‘Larval transport and dispersal in the coastal ocean and consequences for population connectivity’. Oceanography. 20, 3: 22-39.
Pinti, D. L., 2005. ‘The origin and evolution of the oceans’. In: M. Gargaud, B. Barbier, H. Martin & Reisse, J. (eds), Lectures in Astrobiology: Volume I. Berlin, Heidelberg: Springer Berlin Heidelberg, pp. 83-112.
Pitarch, J., 2020. ‘A review of Secchi’s contribution to marine optics and the foundation of Secchi disk science’. Oceanography, 33, 3: 26-37.
Pope, R. M. & Fry, E. S., 1997. ‘Absorption spectrum (380-700 nm) of pure water. II. integrating cavity measurements’. Applied Optics, 36, 33: 8710-23.
Prawiradilaga, D. M. & Irham, M., 2024. ‘Endemism’. In: S. M. Scheiner (ed.). Encyclopedia of Biodiversity. 3rd ed.. Oxford: Academic Press, pp. 72-83.
Puckett, B. J., Theuerkauf, S. J., Eggleston, D. B., Guajardo, R., Hardy, C., Gao, J. & Luettich, R. A., 2018. ‘Integrating larval dispersal, permitting, and logistical factors within a validated habitat suitability index for oyster restoration’. Frontiers in Marine Science, 5, 76.
Rafiudeen, R. A., Anwardeen, N. H., Lavanya, V., Jamal, S. & Ahmed, N., 2024. ‘Coral Reef Metabolites for Cancer Treatment’. Current Pharmacology Reports, 11, 1: 3.
Raghukumar, K., Nelson, T., Jacox, M., Chartrand, C., Fiechter, J., Chang, G., Cheung, L. & Roberts, J., 2023. ‘Projected cross-shore changes in upwelling induced by offshore wind farm development along the california coast’. Communications Earth & Environment, 4, 1: 116.
Randall, J. E., 2005. ‘A Review of mimicry in marine fishes’. Zoological Studies, 44, 3: 299.
Rijke, A. M., 1968. ‘The water repellency and feather structure of cormorants, Phalacrocoracidae’. Journal of Experimental Biology, 48, 1: 185-189.
Roberts, J. M., Wheeler, A. J. & Freiwald, A., 2006. ‘reefs of the deep: the biology and geology of cold-water coral ecosystems’. Science, 312, 5773: 543-547.
Rodríguez-Flores, P. C., Seid, C. A., Rouse, G. W. & Giribet, G., 2023. ‘Cosmopolitan abyssal lineages? a systematic study of east pacific deep-sea squat lobsters (Decapoda: Galatheoidea: Munidopsidae)’. Invertebrate Systematics, 37, 1: 14-60.
Röttgers, R., McKee, D. & Utschig, C., 2014. ‘temperature and salinity correction coefficients for light absorption by water in the visible to infrared spectral region’. Optics Express, 22, 21: 25093-25108.
Ryther, J. H., 1969. ‘Photosynthesis and fish production in the sea’. Science, 166, 3901: 72-76.
Sarafian, A. R., Hauri, E. H., McCubbin, F. M., Lapen, T. J., Berger, E. L., Nielsen, S. G., Marschall, H. R., Gaetani, G. A., Righter, K. & Sarafian, E., 2017. ‘Early accretion of water and volatile elements to the inner Solar System: evidence from angrites’. Philosophical Transactions of the Royal Society A, 375, 2094: 20160209.
Schröder-Ritzrau, A., Freiwald, A. and Mangini, A., 2005. ‘U/Th-dating of deep-water corals from the eastern North Atlantic and the Western Mediterranean Sea’. In: A. Freiwald & Roberts, J. M. (eds). Cold-Water Corals and Ecosystems. Berlin, Heidelberg: Springer Berlin Heidelberg, pp. 157-172.
Seebacher, F., 2009. ‘Responses to temperature variation: integration of thermoregulation and metabolism in vertebrates’. Journal of Experimental Biology, 212, 18: 2885-2891.
Selig, E. R., Casey, K. S. & Bruno, J. F., 2010. ‘New insights into global patterns of ocean temperature anomalies: implications for coral reef health and management’. Global Ecology and Biogeography, 19, 3: 397-411.
Sharma, S., Sharma, V. & Chatterjee, S., 2021. ‘Microplastics in the Mediterranean Sea: sources, pollution intensity, sea health, and regulatory policies’. Frontiers in Marine Science, 8, 634934.
Shiklomanov, I. A., 1990. ‘World freshwater resources’. In: P. H. Gleick (ed.). Water in Crisis: A Guide to the World’s Freshwater Resources. New York: Oxford University Press,pp. 13-24.
Smith, A. P., 2008. ‘Proof of the atmospheric greenhouse effect’. arXiv, arXiv:0802.4324.
Sobha, T. R., Vibija, C. P. & Fahima, P., 2023. ‘Coral reef: a hot spot of marine biodiversity’. In: S. T. Sukumaran & Keerthi, T.R. (eds). Conservation and sustainable utilization of bioresources. Singapore:.Springer Nature Singapore, pp. 171-194.
Srinivasan, S., Chhatre, S. S., Guardado, J. O., Park, K. C., Parker, A. R., Rubner, M. F., McKinley, G. H. & Cohen, R. E., 2014. ‘Quantification of feather structure, wettability and resistance to liquid penetration’. Journal of The Royal Society Interface, 11, 96:20140287.
Stevens, P. E., Allen, V. & Bruck, J. N., 2023. ‘A quieter ocean: experimentally derived differences in attentive responses of tursiops truncatus to anthropogenic noise playbacks before and during the COVID-19-related anthropause’. Animals, 13, 7: 1269.
Stewart, H. A. & Jamieson, A. J., 2019. ‘The five deeps: the location and depth of the deepest place in each of the world’s oceans’. Earth-Science Reviews, 197, 102896.
Stillinger, F. H., 1980. ‘Water revisited’. Science, 209, 4455: 451-457.
Stolarski, J., Kitahara, M. V., Miller, D. J., Cairns, S. D., Mazur, M. & Meibom, A. 2011. ‘The ancient evolutionary origins of scleractinia revealed by azooxanthellate corals’. BMC Evolutionary Biology, 11, 316.
Strang, C. I. and Bosker, T., 2024. ‘The impact of climate change on marine mega-decapod ranges: a systematic literature review’. Fisheries Research, 280, 107165
Trenberth, K., 2011. ‘Changes in precipitation with climate change’. Climate Research, 47, 123-138.
Underwood, C. N., Davies, T. W. & Queirós, A. M. 2017. ‘Artificial light at night alters trophic interactions of intertidal invertebrates’. Journal of Animal Ecology, 86, 4: 781-789.
USGS, 2019. Evaporation and the Water Cycle. U.S. Geological Survey’s (USGS) Water Science School Webpage, viewed: 28 September 2025, <https://www.usgs.gov/water-science-school/science/evaporation-and-water-cycle>.
USGS, 2025. Water Cycle. U.S. Geological Survey’s (USGS) Water Science School Webpage, viewed: 28 September 2025, <https://www.usgs.gov/water-science-school/water-cycle>.
Virgen-Urcelay, A. & Donner, S. D., 2023. ‘Increase in the extent of mass coral bleaching over the past half-century, based on an updated global database’. PLoS One, 18, 2: e0281719.
Vogt-Vincent, N. S., Mitarai, S. & Johnson, H. L., 2023. ‘High-frequency variability dominates potential connectivity between remote coral reefs’. Limnology and Oceanography, 68, 12: 2733-2748.
von Schuckmann, K., Le Traon, P.-Y., Smith, N., Pascual, A., Djavidnia, S., Gattuso, J.-P., Grégoire, M., Nolan, G., Aaboe, S., Aguiar, E., Álvarez Fanjul, E., Alvera-Azcárate, A., Aouf, L., Barciela, R., Behrens, A., Belmonte Rivas, M., Ben Ismail, S., Bentamy, A., Borgini, M., Brando, V. E., Bensoussan, N., Blauw, A., Bryère, P., Buongiorno Nardelli, B., Caballero, A., Çağlar Yumruktepe, V., Cebrian, E., Chiggiato, J., Clementi, E., Corgnati, L., de Alfonso, M., de Pascual Collar, Á., Deshayes, J., Di Lorenzo, E., Dominici, J.-M., Dupouy, C., Drévillon, M., Echevin, V., Eleveld, M., Enserink, L., García Sotillo, M., Garnesson, P., Garrabou, J., Garric, G., Gasparin, F., Gayer, G., Gohin, F., Grandi, A., Griffa, A., Gourrion, J., Hendricks, S., Heuzé, C., Holland, E., Iovino, D., Juza, M., Kurt Kersting, D., Kipson, S., Kizilkaya, Z., Korres, G., Kõuts, M., Lagemaa, P., Lavergne, T., Lavigne, H., Ledoux, J.-B., Legeais, J.-F., Lehodey, P., Linares, C., Liu, Y., Mader, J., Maljutenko, I., Mangin, A., Manso-Narvarte, I., Mantovani, C., Markager, S., Mason, E., Mignot, A., Menna, M., Monier, M., Mourre, B., Müller, M., Nielsen, J. W., Notarstefano, G., Ocaña, O., Pascual, A., Patti, B., Payne, M. R., Peirache, M., Pardo, S., Pérez Gómez, B., Pisano, A., Perruche, C., Peterson, K. A., Pujol, M.-I., Raudsepp, U., Ravdas, M., Raj, R. P., Renshaw, R., Reyes, E., Ricker, R., Rubio, A., Sammartino, M., Santoleri, R., Sathyendranath, S., Schroeder, K., She, J., Sparnocchia, S., Staneva, J., Stoffelen, A., Szekely, T., Tilstone, G. H., Tinker, J., Tintoré, J., Tranchant, B., Uiboupin, R., Van der Zande, D., von Schuckmann, K., Wood, R., Woge Nielsen, J., Zabala, M., Zacharioudaki, A., Zuberer, F. & Zuo, H., 2019. ‘Copernicus marine service ocean state eport, issue 3’. Journal of Operational Oceanography, 12, sup1: S1-S123.
Wagner, T., Schliep, E. M., North, J. S., Kundel, H., Custer, C. A., Ruzich, J. K. & Hansen, G. J. A., 2023. ‘Predicting climate change impacts on poikilotherms using physiologically guided species abundance models’. Proceedings of the National Academy of Sciences, 120, 15: e2214199120.
Weiner, J., 1999. Życie i ewolucja biosfery. Warszawa: Wydawnictwo Naukowe PWN.
Wright, L., 2023. ‘The impact of overfishing on marine ecosystems’. Poultry, Fisheries & Wildlife Sciences, 11, 221.
Wright, S. L., Thompson, R. C. & Galloway, T. S., 2013. ‘The physical impacts of microplastics on marine organisms: a review’. Environmental Pollution, 178, 483-492.
Wu, J., Desch, S. J., Schaefer, L., Elkins-Tanton, L. T., Pahlevan, K. & Buseck, P. R., 2018. ‘Origin of Earth’s water: chondritic inheritance plus nebular ingassing and storage of hydrogen in the core’. Journal of Geophysical Research: Planets, 123, 10: 2691-2712.
Xiu, P., Chai, F., Curchitser, E. N. & Castruccio, F. S., 2018. ‘Future changes in coastal upwelling ecosystems with global warming: the case of the California Current System’. Scientific Reports, 8, 1: 2866.
Videos
SDG 14: life below water. Introduction
Aleksandra Skawina
SDG 14: life below water. Plastic pollution
Aleksandra Skawina
SDG 14: life below water. Fresh water
Aleksandra Skawina
Take a quiz
Test your knowledge on this topic.
Take this short quiz and see if you can score 100%.